When asked to justify the research I do, I always struggle a little to explain my dissatisfaction with the traditional descriptive, non-quantitative explanations of molecular biological systems. As a glance at a classic molecular cell biology textbook will easily demonstrate, molecular biologists have been tremendously successful with verbal or semi-quantitative explanations of what goes on inside the cell. And in any case, the complexity of the cell is extremely daunting for the would-be theoretical biologist. Cells are among the most complex systems we know of - they're made of hundreds of thousands of different types of components, some present at only a few copies per cell, while others can be found at hundreds of millions of copies. Not only are cells extremely heterogeneous in terms of their composition; they're extremely organized.
So when I argue that we need to understand molecular biological systems on a more quantitative level, I'm not knocking traditional molecular biologists, who have been both very clever and spectacularly successful.
In spite of this success, I find most 'blob and arrow' explanations of gene regulatory networks or signal transduction pathways dissatisfying. Why?
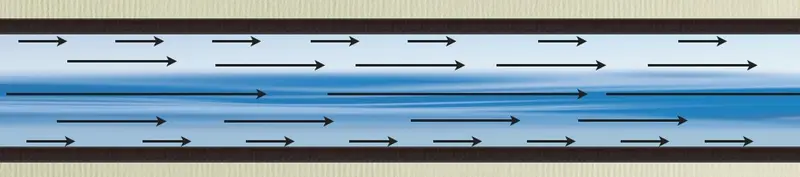 To illustrate my discontent, I'm going to tell you how I think a typical molecular biologist would explain the laminar flow of water through a pipe:
To illustrate my discontent, I'm going to tell you how I think a typical molecular biologist would explain the laminar flow of water through a pipe:
In answer to the question, how does water flow through a pipe?, a molecular biologist would do a series of clever experiments and come up with the observation that, under certain conditions, water flows through a pipe in layers, with each layer moving at a different velocity. Water on the outside of the flow, adjacent to the pipe walls, moves most slowly, while water towards the center flows most rapidly. As the overall flow rate is increased past a certain point, this organized flow deteriorates into something more chaotic. Our hypothetical biologist would probably also invoke friction as a primary cause of the structure of laminar flow - water molecules adjacent to the pipe walls experience more friction than water molecules in the center of the flow.
That would be pretty much it. Completely absent would be any quantitative, physically-based explanation for why you get the observed distribution of velocities at a given flow rate, or how you would expect those velocities to change as you alter the flow rate or change the composition of the pipe walls. An idea like the Reynolds Number would be much too abstract.
This is the level of most explanations in molecular cell biology - you can see this as you read through any good textbook on molecular biology, which typically contains primarily descriptive paragraphs similar to what I just wrote about laminar flow (although the writing is the textbooks is much better than mine).
Here is a passage on how specificity of signal transmission is achieved in a classic MAP Kinase pathway (from Lodish, section 20.5 - see the link to the book above):
To illustrate one way multiple MAP kinases are segregated, we consider how a change in osmolarity activates Ste11 but does not lead to activation of downstream components in the mating pathway. There are two osmoregulatory MAP kinase pathways in S. cerevisiae; both lead to activation of the MAP kinase Hog1, but only one pathway requires Ste11. The dual-specificity MEK in this osmoregulatory pathway, called Pbs2, also functions as a scaffold for assembly of a large signaling complex. Pbs2 binds to Hog1, Ste11, and Sho1 (the osmolarity-sensitive receptor). Transmission of the signal from Sho1 to Hog1 occurs within the complex assembled by Pbs2. Recall that in the mating pathway, the scaffold protein Ste5 likewise stabilizes a large complex including Ste11. In both cases, the common component Ste11 is constrained within a large complex that forms in response to a specific extracellular signal, and signaling downstream from Ste11 is restricted to the complex containing it (Figure 20-33). As a result, exposure of yeast cells to mating factors induces activation of a single MAP kinase, Fus3, while exposure to a high osmolarity induces activation only of Hog1.
It just goes on like this, and it's not the textbook writers' fault. This is how molecular biology answers questions.
If engineers were limited to that kind of understanding of laminar flow, they'd have a difficult time designing airplane wings and car engines. And frankly, it's much more satisfying to explain the behavior of flowing water as a phenomenon that emerges from underlying general principles or the physical properties of water. To get at that kind of explanation, you have to be quantitative, you have to abstract key elements of the system you're studying, and you have to come up with equations that rigorously describe those abstractions.
Naturally, this is much easier to talk about than to actually accomplish. Like I said, biology is more complex than just about any other natural system that scientists study.
Yet even though developing more principles-based, quantitative explanations in molecular biology is difficult, it's not difficult to see why our current explanations are unsatisfying.
My interest in science was always essentially limited to the study of principles, which best explains my conduct in its entirety. That I have published so little is attributable to the same circumstance, for the burning desire to grasp principles has caused me to spend most of my time on fruitless endeavors.
- Albert Einstein, quoted in Einstein's Clocks, Poincaré's Maps, by Peter Galison, p. 241
Read the feed:




